Porcine TGF-beta 1 Protein
Discontinued Product
101-B1 has been discontinued and is replaced by 7754-BH.
Porcine TGF-beta 1 Protein Summary
Product Specifications
Product Datasheets
101-B1/CF (carrier free)
Discontinued Product
101-B1
| Formulation | Lyophilized from a 0.2 μm filtered solution in Acetonitrile and TFA with BSA as a carrier protein. |
| Reconstitution | Reconstitute at 10 μg/mL in sterile 4 mM HCl containing at least 0.1% human or bovine serum albumin. |
| Shipping | The product is shipped at ambient temperature. Upon receipt, store it immediately at the temperature recommended below. |
| Stability & Storage: | Use a manual defrost freezer and avoid repeated freeze-thaw cycles.
|
Reconstitution Calculator
Background: TGF-beta 1
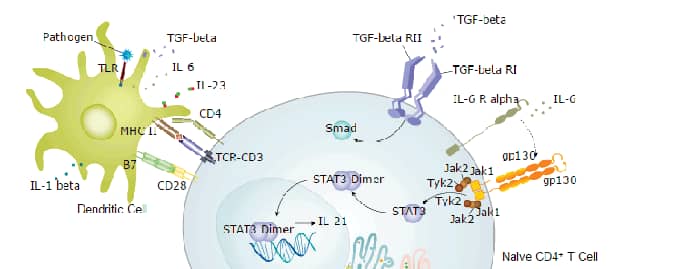
TGF- beta 1 (transforming growth factor beta 1) is one of three closely related mammalian members of the large TGF-beta superfamily that share a characteristic cystine knot structure (1 - 7). TGF-beta 1, -2 and -3 are highly pleiotropic cytokines that are proposed to act as cellular switches that regulate processes such as immune function, proliferation and epithelial-mesenchymal transition (1 - 4). Each TGF-beta isoform has some non redundant functions; for TGF-beta 1, mice with targeted deletion show defects in hematopoiesis and endothelial differentiation, and die of overwhelming inflammation (2). Porcine TGF-beta 1 cDNA encodes a 390 amino acid (aa) precursor that contains a 29 aa signal peptide and a 361 aa proprotein (8). A furin-like convertase processes the proprotein to generate an N-terminal 220 aa latency-associated peptide (LAP) and a C-terminal 112 aa mature TGF- beta 1 (8, 9). Disulfide-linked homodimers of LAP and TGF-beta 1 remain non-covalently associated after secretion, forming the small latent TGF-beta 1 complex (8 - 10). Covalent linkage of LAP to one of three latent TGF-beta binding proteins (LTBPs) creates a large latent complex that may interact with the extracellular matrix (9, 10). TGF-beta is activated from latency by pathways that include actions of the protease plasmin, matrix metalloproteases, thrombospondin 1 and a subset of integrins (10). Mature porcine TGF-beta 1 shows 100% aa identity with human, dog and cow TGF-beta 1 and 99% aa identity with mouse, rat and horse TGF-beta 1. It demonstrates cross species activity (1). TGF-beta 1 signaling begins with high-affinity binding to a type II ser/thr kinase receptor termed TGF-beta RII. This receptor then phosphorylates and activates a second ser/thr kinase receptor, TGF-beta RI (also called activin receptor-like kinase (ALK) -5), or alternatively, ALK-1.This complex phosphorylates and activates Smad proteins that regulate transcription (3, 11, 12). Contributions of the accessory receptors betaglycan (also known as TGF-beta RIII) and endoglin, or use of Smad-independent signaling pathways, allow for disparate actions observed in response to TGF-beta in different contexts (11).
- Sporn, M.B. (2006) Cytokine Growth Factor Rev. 17:3.
- Dunker, N. and K. Krieglstein (2000) Eur. J. Biochem. 267:6982.
- Wahl, S.M. (2006) Immunol. Rev. 213:213.
- Chang, H. et al. (2002) Endocr. Rev. 23:787.
- Lin, J.S. et al. (2006) Reproduction 132:179.
- Hinck, A.P. et al. (1996) Biochemistry 35:8517.
- Mittl, P.R.E. et al. (1996) Protein Sci. 5:1261.
- Kondaiah, P. et al. (1988) J. Biol. Chem. 263:18313.
- Miyazono, K. et al. (1988) J. Biol. Chem. 263:6407.
- Oklu, R. and R. Hesketh (2000) Biochem. J. 352:601.
- de Caestecker, M. et al. (2004) Cytokine Growth Factor Rev. 15:1.
- Zuniga, J.E. et al. (2005) J. Mol. Biol. 354:1052.
Citations for Porcine TGF-beta 1 Protein
R&D Systems personnel manually curate a database that contains references using R&D Systems products. The data collected includes not only links to publications in PubMed, but also provides information about sample types, species, and experimental conditions.
22
Citations: Showing 1 - 10
Filter your results:
Filter by:
-
Angiotensin-II-Evoked Ca2+ Entry in Murine Cardiac Fibroblasts Does Not Depend on TRPC Channels
Authors: JE Camacho Lo, A Marx, AE Kraft, A Schürger, C Richter, A Dietrich, P Lipp, L Birnbaumer, M Freichel
Cells, 2020-01-29;9(2):.
Species: Mouse
Sample Types: Whole Cells
Applications: Bioassay -
Regulation of fibroblast Fas expression by soluble and mechanical pro-fibrotic stimuli
Authors: AE Dodi, IO Ajayi, C Chang, M Beard, SL Ashley, SK Huang, VJ Thannickal, DJ Tschumperl, TH Sisson, JC Horowitz
Respir. Res., 2018-05-10;19(1):91.
Species: Human
Sample Types: Whole Cells
Applications: Bioassay -
Specialized fibroblast differentiated states underlie scar formation in the infarcted mouse heart
Authors: X Fu, H Khalil, O Kanisicak, JG Boyer, RJ Vagnozzi, BD Maliken, MA Sargent, V Prasad, I Valiente-A, BC Blaxall, JD Molkentin
J. Clin. Invest., 2018-04-16;0(0):.
Species: Mouse
Sample Types: Whole Cells
Applications: Bioassay -
CD96 expression determines the inflammatory potential of IL-9-producing Th9 cells
Authors: K Stanko, C Iwert, C Appelt, K Vogt, J Schumann, FJ Strunk, S Ahrlich, S Schlickeis, C Romagnani, K Jürchott, C Meisel, G Willimsky, AA Kühl, B Sawitzki
Proc. Natl. Acad. Sci. U.S.A., 2018-03-12;0(0):.
Species: Human
Sample Types: Whole Cells
Applications: Bioassay -
SMAD4 feedback regulates the canonical TGF-? signaling pathway to control granulosa cell apoptosis
Authors: X Du, Z Pan, Q Li, H Liu, Q Li
Cell Death Dis, 2018-02-02;9(2):151.
Species: Porcine
Sample Types: Whole Cells
Applications: Bioassay -
Diverse functions of clusterin promote and protect against the development of pulmonary fibrosis
Authors: L Peix, IC Evans, DR Pearce, JK Simpson, TM Maher, RJ McAnulty
Sci Rep, 2018-01-30;8(1):1906.
Species: Human
Sample Types: Whole Cells
Applications: Bioassay -
Glutaminolysis is required for TGF-?1-induced myofibroblast differentiation and activation
Authors: K Bernard, NJ Logsdon, GA Benavides, Y Sanders, J Zhang, VM Darley-Usm, VJ Thannickal
J. Biol. Chem., 2017-12-08;0(0):.
Species: Human
Sample Types: Whole Cells
Applications: Bioassay -
Fibroblast-specific TGF-?-Smad2/3 signaling underlies cardiac fibrosis
Authors: H Khalil, O Kanisicak, V Prasad, RN Correll, X Fu, T Schips, RJ Vagnozzi, R Liu, T Huynh, SJ Lee, J Karch, JD Molkentin
J. Clin. Invest., 2017-09-11;0(0):.
Species: Mouse
Sample Types: Whole Cells
Applications: Bioassay -
Low doses of cholera toxin and its mediator cAMP induce CTLA-2 secretion by dendritic cells to enhance regulatory T cell conversion
Authors: C Silva-Vilc, K Pletinckx, M Lohnert, V Pavlovic, D Ashour, V John, E Vendelova, S Kneitz, J Zhou, R Chen, T Reinheckel, TD Mueller, J Bodem, MB Lutz
PLoS ONE, 2017-07-31;12(7):e0178114.
Species: Mouse
Sample Types: Whole Cells
Applications: Bioassay -
TGF-? signaling controls FSHR signaling-reduced ovarian granulosa cell apoptosis through the SMAD4/miR-143 axis
Cell Death Dis, 2016-11-24;7(11):e2476.
Species: Porcine
Sample Types: Whole Cells
Applications: Bioassay -
Context-dependent switch in chemo/mechanotransduction via multilevel crosstalk among cytoskeleton-regulated MRTF and TAZ and TGF?-regulated Smad3
Nat Commun, 2016-05-18;7(0):11642.
Species: Porcine
Sample Types: Whole Cells
Applications: Bioassay -
Serotonin 2B Receptor Antagonism Prevents Heritable Pulmonary Arterial Hypertension.
Authors: West J, Carrier E, Bloodworth N, Schroer A, Chen P, Ryzhova L, Gladson S, Shay S, Hutcheson J, Merryman W
PLoS ONE, 2016-02-10;11(2):e0148657.
Species: Mouse
Sample Types: Whole Cells
Applications: Bioassay -
Metabolic Reprogramming Is Required for Myofibroblast Contractility and Differentiation.
Authors: Bernard K, Logsdon N, Ravi S, Xie N, Persons B, Rangarajan S, Zmijewski J, Mitra K, Liu G, Darley-Usmar V, Thannickal V
J Biol Chem, 2015-08-28;290(42):25427-38.
Species: Human
Sample Types: Whole Cells
Applications: Bioassay -
IkappaBNS regulates murine Th17 differentiation during gut inflammation and infection.
Authors: Annemann M, Wang Z, Plaza-Sirvent C, Glauben R, Schuster M, Ewald Sander F, Mamareli P, Kuhl A, Siegmund B, Lochner M, Schmitz I
J Immunol, 2015-02-18;194(6):2888-98.
Species: Mouse
Sample Types: Whole Cells
Applications: Bioassay -
SMAD-independent down-regulation of caveolin-1 by TGF-beta: effects on proliferation and survival of myofibroblasts.
Authors: Sanders Y, Cui Z, Le Saux C, Horowitz J, Rangarajan S, Kurundkar A, Antony V, Thannickal V
PLoS ONE, 2015-02-06;10(2):e0116995.
Species: Human
Sample Types: Whole Cells
Applications: Bioassay -
RGD surface functionalization of the hydrophilic acrylic intraocular lens material to control posterior capsular opacification.
Authors: Huang Y, Bertrand V, Bozukova D, Pagnoulle C, Labrugere C, De Pauw E, De Pauw-Gillet M, Durrieu M
PLoS ONE, 2014-12-11;9(12):e114973.
Species: Porcine
Sample Types: Whole Cells
Applications: Bioassay -
Tc17, a unique subset of CD8 T cells that can protect against lethal influenza challenge.
Authors: Hamada H, Garcia-Hernandez Mde L, Reome JB, Misra SK, Strutt TM, McKinstry KK, Cooper AM, Swain SL, Dutton RW
J. Immunol., 2009-03-15;182(6):3469-81.
Species: Mouse
Sample Types: Whole Cells
Applications: Bioassay -
TH17 cells mediate steroid-resistant airway inflammation and airway hyperresponsiveness in mice.
Authors: McKinley L, Alcorn JF, Peterson A, Dupont RB, Kapadia S, Logar A, Henry A, Irvin CG, Piganelli JD, Ray A, Kolls JK
J. Immunol., 2008-09-15;181(6):4089-97.
Species: Mouse
Sample Types: Whole Cells
Applications: Bioassay -
The aberrant methylation of TSP1 suppresses TGF-beta1 activation in colorectal cancer.
Authors: Rojas A, Meherem S, Kim YH, Washington MK, Willis JE, Markowitz SD, Grady WM
Int. J. Cancer, 2008-07-01;123(1):14-21.
Species: Human
Sample Types: Recombinant Protein
Applications: Bioassay -
3-Phosphoinositide-dependent PDK1 negatively regulates transforming growth factor-beta-induced signaling in a kinase-dependent manner through physical interaction with Smad proteins.
Authors: Seong HA, Jung H, Kim KT, Ha H
J. Biol. Chem., 2007-02-27;282(16):12272-89.
Species: Human
Sample Types: Whole Cells
Applications: Bioassay -
Transforming growth factor beta1 regulates follistatin mRNA expression during in vitro bovine granulosa cell differentiation.
Authors: Fazzini M, Vallejo G, Colman-Lerner A, Trigo R, Campo S, Baranao JL, Saragueta PE
J. Cell. Physiol., 2006-04-01;207(1):40-8.
Species: Bovine
Sample Types: Whole Cells
Applications: Bioassay -
Xenopus neurula left-right asymmetry is respeficied by microinjecting TGF-beta5 protein.
Authors: Mogi K, Goto M, Ohno E, Azumi Y, Takeuchi S, Toyoizumi R
Int. J. Dev. Biol., 2003-02-01;47(1):15-29.
Species: Xenopus
Sample Types: In Vivo
Applications: In Vivo
FAQs
No product specific FAQs exist for this product, however you may
View all Proteins and Enzyme FAQsReviews for Porcine TGF-beta 1 Protein
There are currently no reviews for this product. Be the first to review Porcine TGF-beta 1 Protein and earn rewards!
Have you used Porcine TGF-beta 1 Protein?
Submit a review and receive an Amazon gift card.
$25/€18/£15/$25CAN/¥75 Yuan/¥2500 Yen for a review with an image
$10/€7/£6/$10 CAD/¥70 Yuan/¥1110 Yen for a review without an image